Введите слово или словосочетание на любом языке 👆
Язык:
Перевод и анализ слов искусственным интеллектом ChatGPT
На этой странице Вы можете получить подробный анализ слова или словосочетания, произведенный с помощью лучшей на сегодняшний день технологии искусственного интеллекта:
- как употребляется слово
- частота употребления
- используется оно чаще в устной или письменной речи
- варианты перевода слова
- примеры употребления (несколько фраз с переводом)
- этимология
Что (кто) такое Зародыш - определение
СТРАНИЦА ЗНАЧЕНИЙ
Зародыш
1) у животных и человека З., или эмбрион, - организм в раннем периоде развития - от оплодотворённого яйца (зиготы (См. Зигота)) до появления способности к самостоятельному питанию и активному передвижению. Развитие З. происходит, как правило, в яйцевых оболочках или специальных органах материнского организма. См. Зародышевое развитие, Зародышевые оболочки.
2) У растений З., или эмбрио, - зачаток новой особи, развивающийся из зиготы, которая возникает в результате оплодотворения яйцеклетки зародышевого мешка (См. Зародышевый мешок) или из клеток нуцеллуса или интегумента (в случае Апомиксиса). Наиболее простой тип эмбрионального развития у зелёных водорослей, например у спирогиры, зигота которой не образует спорофитной нити, а сразу приступает к Мейозу. У бурых водорослей, например у фукуса, зигота прикрепляется к субстрату, делится, после чего образует эмбриональное тело с точкой роста Таллома (Рис.1). У мохообразных З. прорастает на материнском организме - Гаметофите, за счёт которого он питается, затем образует спорогенную ткань, где формируются споры (Рис.2). Такой тип З. не встречается среди др. групп высших растений. У древнейших высших растений - псилотовых - развитие З. протекает в ткани гаметофита, где происходит ряд последовательных делений зиготы. Из клеток нижней части формируется ножка, внедряющаяся в ткань гаметофита, в верхней - закладывается точка роста побега с двумя меристематическими бугорками (см. Меристема). З. здесь - поляризованная структура, один конец которой образует наземный побег, другой - корневище (Рис.3). У плауновидных З. имеет два симметричных бугорка будущих зародышевых листочков, между которыми находится точка роста в виде верхушечной клетки (Рис.4). Развитие З. у хвощей характеризуется правильным планом сегментации в виде 4 зон, дающих начало побегу, корню, первому листу и ножке (Рис.5). У папоротниковидных в первых 4 клетках, возникших в результате двух первых делении зиготы, определено место закладки верхушки побега, первого листа, корня и ножки (Рис.6). У голосеменных начальный цикл развития З. характеризуется свободноядерной, или ценоцитной, стадией (у саговников, например, свободных ядер 1024), которая сменяется образованием клеток; затем в формирующемся З. происходит закладка точек роста побега, корня и семядольных бугорков (у некоторых видов сосны - до 18) (Рис.7). У большинства покрытосеменных зигота делится на две клетки: апикальную, дающую начало собственно З., п базальную - подвеску, или суспензору.
В Эмбриогенезе различают 2 фазы развития: предзародышевую (от зиготы до образования эмбриодермы) и зародышевую (закладка основных структур: точки роста побега, семядоли, гипокотиля и центрального зародышевого корешка). На основе первых этапов деления предзародыша были созданы классификации эмбриональных типов, используемые систематиками при решении вопросов эволюции и филогении растений. Предзародышевая стадия развития двудольных и однодольных одинакова. На более поздних стадиях у однодольных апикальная зона проэмбрио даёт начало только 1 семядольному бугорку; у двудольных таких бугорков может возникнуть 2, а у некоторых видов 3 и даже больше, что приводит к образованию многосемядольности. У покрытосеменных растений развитие З., в отличие от голосеменных, сопровождается образованием клеточных перегородок (Рис.8,9). Лишь у пионов обнаружен новый тип эмбриогенеза, при котором в зиготе вначале возникают свободные ядра, а затем такая многоядерная структура становится клеточной и на её периферии возникают проэмбриональные бугорки; из них в собственно З. развивается только один. По наличию или отсутствию у З. покрытосеменных зелёного пигмента - хлорофилла - они делятся на 2 группы: хлороэмбриофиты (бобовые, крестоцветные и др.) илейкоэмбриофиты (лютиковые, злаковые и др.). В процессе эволюции среди некоторых групп покрытосеменных, например у растений паразитов (заразиховые) и сапрофитов (орхидные), наблюдается -редукция З. Источником питания З. служит Эндосперм, который у одних видов (бобовые) поглощается целиком растущим З., у др. (злаки) - сохраняется. Иногда З. в семени окружен не только эндоспермом, но и др. запасной тканью - периспермом, развивающимся из клеток нуцеллуса (свёкла, чёрный перец и др.).
Лит.: Баранов П. А., История эмбриологии растений в связи с развитием представлений о зарождении организмов, М.- Л., 1955; Яковлев М. С., Эмбриогенез и его значение для филогении растений, М. - Л., 1960; Поддубная-Арнольди В. А., Общая эмбриология покрытосеменных растений, М., 1964; Тахтаджян А. Л., Основы эволюционной морфологии покрытосеменных, М. - Л., 1964.
М. С. Яковлев.
Рис.1. Развитие зародыша у растений разных систематических групп. У бурой водоросли фукус (а - г).
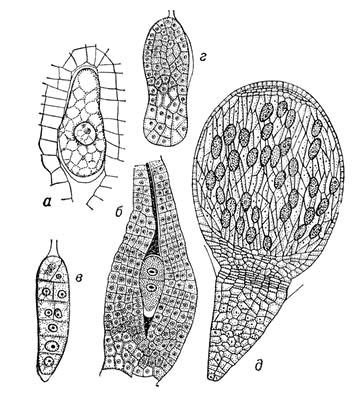
Рис.2. Развитие зародыша у растений разных систематических групп. У мохообразных (Conocephalum conicum): a - зигота, б - первое деление зиготы горизонтальной перегородкой, в - образование поперечных перегородок, г - апикальный и базальный концы зародыша, дифференциация археспориальной ткани, д - зрелый зародыш, ставший спорогонием.
Рис.3. Развитие зародыша у растений разных систематических групп. У псилотовых (Tmesipteris tannensis): а - первое деление зиготы, б и в - проэмбрио, г - закладка апикальных клеток, д - начало образования дихотомически разветвленного зародыша.
Рис.4. Развитие зародыша у растений разных систематических групп. У плауновидных (Selaginella martensii): а, б, в - закладка апикальной точки роста (заштриховано), г - зрелый зародыш с прокамбиальным тяжом, точкой роста побега и листовыми зачатками, д - зародыш в ткани гаметофита.
Рис.5. Развитие зародыша у растений разных систематических групп. У хвощей (Equisetum debileu Е. arwense): а и б - начальные фазы эмбриогенеза, в - более поздняя фаза, в апикальной части видна верхушечная клетка стебля.
Рис.6. Развитие зародыша у растений разных систематических групп. У папоротника (Onoklea sp.): зрелый зародыш, хорошо развит первый лист (а) и корешок (б), закладывается второй лист (в), виден прокамбиальный тяж (г), дающий начало сосудистой системе, справа ножка (д), через которую в зародыш поступают питательные вещества из ткани гаметофита, или заростка.
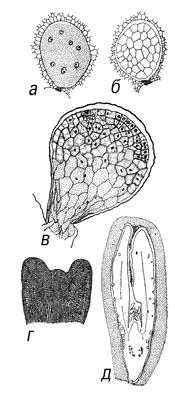
Рис.7. Развитие зародыша у растений разных систематических групп. У голосеменных (Ginkgo biloba): а - проэмбрио в свободноядерной фазе, б - клеткообразование, в - проэмбрио, начало дифференциации клеток апикальной зоны, г - появление семядольных бугорков и точки роста, д - зрелый, хорошо дифференцированный зародыш.
Рис.8. Зародыш растений: 1 - зародыш (а) в семени кедровой сосны; 2-8 - различные стадии развития зародыша у некоторых видов табака: 2 - оплодотворённые яйцеклетка и центральная клетка зародышевого мешка, 3 - двухклеточный предзародыш, 4 - четырёхклеточный предзародыш, 5-6 - многоклеточные предзародыши с подвесками и эндоспермом.
Рис.9.: Зародыш растений: 7 - начало дифференциации зародыша (закладываются бугорки семядолей), 8 - дифференцированный зародыш, окруженный эндоспермом; у зародыша имеются две семядоли, зачатки стебля и корня, точки роста стебля и корня, корневой чехлик; 9-10 - продольный разрез через семя и зародыш овса Avena sativa; зародыш дифференцирован и имеет: а - одну семядолю (щиток), б - влагалище (колеоптиле), в - первые листочки (перышко), г - точку роста стебля, д - зачатки стебля и е - корня, ж - корневой чехлик, з - колеоризу, и - эпибласт; 11 - семя с недифференцированным зародышем у орхидеи Lycaste skinneri; 12 - зародыш с двумя складчатыми семядолями у хлопчатника Gossypium herbaceum.
Зародыш
Зародыш (медиц.) может в полости матки подвергнуться различного родазаболеваниям. Последние могут быть вызваны аномалиями развития, которыемогут зависеть, как от неправильного эмбрионального анатомическогорасположения, так и от остановки в развитии или уродливого развитияплода вследствие аномалии его придатков. От этих болезней З. следуетотличать такие, которые переходят от матери на плод или являютсянаследственными, равно как и такие, которые развиваются у плодасамостоятельно (идиопатические). Среди болезней З., которые являютсяследствием материнских заболеваний, первое место занимают острые сыпи.Последние всего чаще переходят на плод при оспе, затем при скарлатине,кори и роже. Плод может или вовсе не подвергнуться заражению, или дозаражения умереть от повышения температуры, или, наконец, он заражается;в последнем случае он может родиться к нормальному концу беременности спризнаками проделанной болезни (оспенные рубцы), или же преждевременново время болезни, или, наконец, он может родиться преждевременно,по-видимому, здоровым, но заболеть вскоре после рождения. Из другихинфекционных болезней: сыпной и возвратный тифы, холера и малярия могуттакже переходить на плод. Самую же частую болезнь З. составляет сифилисплода. - Между идиопатическими болезнями З. следует назвать врожденныйрахит, причины которого неизвестны. Гораздо чаще встречается и болееважное значение для родоразрешения имеет водянка головы. которая должнабыть поставлена в зависимость от водянки мозговых желудочков споследовательным расширением черепной покрышки. К этой же группеболезней З. относятся случаи свободной жидкости в животе (ascites),опухоли печени, селезенки, яичка, аневризма аорты, зародышевые кисты.Хирургические заболевания плода могут произойти от внешних повреждений.Последним подвергаются по большей части голова и конечности. Этиповреждения ведут в большинстве случаев к смерти плода и кпреждевременному изгнанию его. Чисто хирургическую болезнь З.представляет нередко встречающийся врожденный вывих тазобедренногосустава.
ЗАРОДЫШ
1. У человека и животных, а также у высших семенных растений: организм на ранней ступени развития, живущий за счет материнского организма либо питательных веществ в яйцеклетке.
2. первое появление, заточное состояние чего-нибудь.
Подавить дурную привычку в зародыше.
Википедия
Зародыш
Заро́дыш:
- Зародыш животных — то же, что эмбрион.
- Зародыш растений — см. зародыш (ботаника)
- Фамилия
- Зародыш, Пётр Иванович (1932—2022) — советский передовик сельскохозяйственного производства, Герой Социалистического Труда (1971).

Примеры употребления для Зародыш
1. Возникает вопрос: считать ли зародыш живым существом?
2. - Внутри него - масса, напоминающая засохший зародыш.
3. Это зародыш искусства и понимание искусства ребенком.
4. Теперь зародыш хранится в районной больнице Ачита.
5. Стволовые клетки человека внедряют в зародыш овцы.